The legend of civilian heroes can always be heard in life. In fact, there are many unsung heroes in the living body to maintain the health of our bodies and work in obscurity all the time. KLHL24 is one of them.
Lead
Under normal conditions, KLHL24, which is a ubiquitin ligase, leads the prey-KRT14 to the abyss (proteasome) without going back, while at the same time maintaining its own skin barrier (self-ubiquitination). Unfortunately, the world is impermanent, genetic mutation (Klhl24-∆N28) What can you do? The Klhl24, which lacks the N-terminal 28 amino acids, seems to be in madness. The self-disciplined self-destruction process is no longer the endless endless elimination of prey, which is the ultimate goal, causing a devastating blow to the cells and tissues that have been vowed to protect.
Epidermolysis bullosa ( EB ) is a skin disease . Common symptoms are blisters and blood blisters and erosions after a slight friction or collision. It occurs in the extremities and the extremities of the extremities. Can involve any part of the body. The main type of hereditary EB is simple EB (simplex EB, EBS), the skin lesions are the most superficial, and generally no scars are left after the healing. Damage to skin structural proteins (such as keratin) can cause skin fragility and trigger EB.
The authors used the analysis of gene mutations in EBS patients as an entry point to explain step by step how KLHL24 dominant mutations cause EB.
Culprit - genetic mutation
The KLHL24 gene mutation was found by sequencing all the exomes of different EB patients' genes (Fig. 1a). It was proved by RT-PCR that KLHL24 is widely expressed in various tissues of the human body, including skin (Fig. 1b).
KLHL24 belongs to the Kelch-like gene family, which encodes a protein with a highly conserved BTB domain and BACK domain that interact with CUL3 and RBX1 to form the CUL3–RBX1–Kelch ubiquitin ligase complex. The Kelch-like protein also has a Kelch domain that is used to recruit substrates that determine the specificity of the E3 ligase.
The KLHL24 mutation is located in the initiation codon region, so translation cannot start normally, and it is hypothesized that Met29 may serve as another translation initiation site. The transfected cells carrying the normal KLHL24 and KLHL24-∆N28 plasmids were separately lysed for immunoblotting, and the corresponding bands of the target protein were found (KLHL24-∆N28; Fig. 1c).
Further verification of the KLHL24-∆N28 translation initiation site, Met29 codon mutation (ATG-ATA) in the case of Met1 mutation (ATA), finally found that KLHL24 expression was inhibited (Fig. 1d), thus demonstrating that Met29 can be used as Another translation start site .
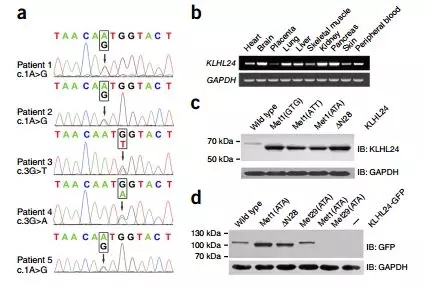
Fig.1 KLHL24 start-codon mutations identified in the five patients result in an N-terminally truncated protein. (a) Sequence chromatograms showing heterozygous c.1A>G (patients 1, 2 and 5), c.3G>T (patient 3) and c.3G>A (patient 4) mutations, affecting the start codon. (b) Expression profile of human KLHL24 determined by RT–PCR, showing ubiquitous expression in representative adult tissues, including skin. (c) Expression of KLHL24 Protein analyzed by immunoblotting in 293T cells transfected with the indicated constructs. (d) Expression of KLHL24 proteins analyzed by immunoblotting in 293T cells transfected with the indicated C-terminally GFP-tagged constructs and untransfected cells (–).
Overexpression of KLHL24 and KLHL24-∆N28 in HaCaT (immortalized human keratinocyte cell line) cells revealed that the expression of KLHL24-∆N28 was much higher than that of normal KLHL24 (Fig. 2a). The guess may be due to protein stability. Sexual differences result in the fact that mRNA levels are not related to the difference in expression of this protein (Fig. 3).
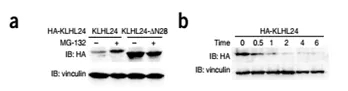

Fig 2. (a) Stably expressed KLHL24-N28 shows markedly higher protein levels than the wild-type protein. MG-132 treatment for 8h increases the protein levels of KLHL24 but not those of KLHL24-∆N28. (bc) Measurement of the Stability of wild-type and mutant KLHL24-∆N28 proteins by cycloheximide (CHX) chase assays.
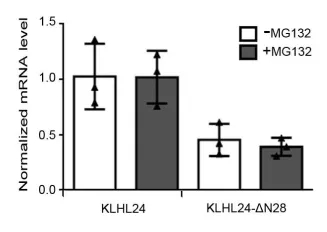
Fig 3. Proteasome inhibitor MG132 treatment increased did not change the mRNA levels of KLHL24 in HaCaT cells stable expressing KLHL24 or KLHL-ΔN28. The mRNA levels were normalized to the mRNA level of KLHL24 in HaCaT cells expressing KLHL24 not treated by MG132
Since it is speculated that this difference in expression is related to protein stability, what happens if a protease inhibitor is added?
Under the action of the protease inhibitor MG-132, the abundance of KLHL24 was greatly improved, but there was no significant change in KLHL24-∆N28 (Fig. 2a), and the half-life of mutant KLHL24-∆N28 (t1/2 = ~4.6 h). It was also measured far beyond the normal KLHL24 (t1/2 = ~0.4 h) (Fig. 2b, c), indicating that KLHL24-∆N28 is more stable than the wild type .
It’s strange to say that there’s a lot less, but life is like opening a hang to directly upgrade King Kong’s body? Why is this?
Combined with previous studies, most E3 ligases are regulated by ubiquitination, so it is speculated that the difference in stability between mutant and normal KLHL24 may also be related to the self-ubiquitination process.
The ubiquitination level of KLHL24 was detected in 293T/HaCaT transfected cells expressing CUL3, RBX1 and KLHL24, and it was found that KLHL24-∆N28 was lower than normal KLHL24 ubiquitination level;
MLN4924 can increase the abundance of KLHL24 by inhibiting ubiquitination and the mutation remains unchanged.
Co-immunoprecipitation experiments showed that the binding ability of KLHL24 and KLHL24-∆N28 to CUL3 was similar, and the stability enhancement of KLHL24-∆N28 was ruled out due to the possibility of losing the ability to bind to CUL3.
The dimerization of Kelch-like protein affects its ubiquitination ability, while both GST and co-immunoprecipitation experiments indicate that KLHL24-∆N28 dimerization ability is not affected.
All the results indicated that the mutant KLHL24-∆N28 lost its self-ubiquitination ability, thereby improving the stability and abundance of the protein.
After all, as a ubiquitin ligase, it is also the duty to direct the substrate to Huang Quan. The time of intimate contact is so short, how can we confirm the right TA from the "sentences of all beings"?

Step 1: Confirm candidate substrates
The KLHL24 Kelch domain (Kelch domain is responsible for binding to the substrate) and the GST fusion protein were expressed separately, and at the same time, Kelch domain mutant (Kelchmut) was used as a parallel control to exclude non-specific binding proteins. The HaCaT cell lysate was sequentially subjected to GST pulldown and mass spectrometry, and some candidate substrates were confirmed. Preliminary mass spectrometry analysis (Fig. 4c) showed that KRT14 is unique, and it is likely to be a substrate for KLHL24.

Further comparison with the control group revealed that KRT14 could bind to KLHL24, KLHL24-∆N28 (Fig. 4c, d, e), while Kelchmut had a significantly lower affinity with KRT14.
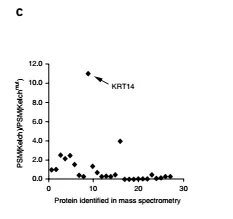
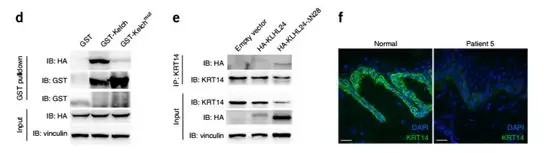

Fig. (c) PSM (peptide spectrum match) score ratios for the proteins identified by mass spectrometry. (d) GST pulldown experiments indicate that GST-Kelch interacts with HA-KRT14. The two GST blots were cropped from the full-length blot (e) Coimmunoprecipitation experiments in HaCaT cells indicate that KRT14 interacts with KLHL24 and KLHL24-∆N28. (f) Immunofluorescent staining shows markedly reduced KRT14 levels in the basal keratinocytes of skin from patient 5 in contrast to normal skin (g) Immunoblotting of skin samples shows a marked reduction in the amount of KRT14 in patient 5 in comparison to normal controls.
Step 2: Verify the substrate
The KRT14 protein content was first determined. The results showed that the KRT14 content in the skin samples of the two patients was significantly lower than that of the age-matched non-sick control group (Fig. 4f, g). At the same time, the content of Keratin 5 (KRT5) which formed a heterodimer with KRT14 was also lower than that of the control group, but the content of keratin 1 (KRT1) did not change (does not form a dimer with KRT14 or KRT5).
Keratin immunofluorescence and skin differentiation markers indicate a significant reduction in keratin in the basal cells of the patient, but the abundance of basal keratin and differentiation markers remains unchanged (Fig. 5).

Figure 5. IF staining of pan-keratin, KRT1, keratin10 (KRT10) and loricrin on skin sections of Patient 5 and a normal control.
Overexpression of KLHL24 in HaCaT cells reduced KRT14 protein levels to a small extent, while overexpression of KLHL24-∆N28 caused a sharp decrease in KRT14 content ("input" in Fig. 4e); half-life test results indicated that KLHL24 and KLHL24-∆N28 Overexpression can reduce the stability of KRT14.
RNAi indicates that down-regulation of KLHL24 increases the level of KRT14 protein, whereas for KLHL24-∆N28, KRT14 content is increased when the siRNA target is located in the coding region, and there is no significant change in the 5' or 3'UTR region.
Further detection of KRT14 ubiquitination level in 293T cells, observed that KLHL24 or KLHL24-∆N28 transient expression combined with CUL3 and RBX1 can increase the ubiquitination level of KRT14.
Based on the above results of genetic analysis and biochemical experiments, KLHL24 acts as an E3 ubiquitin ligase on the substrate KRT14, and its function-acquired mutant KLHL24-∆N28 causes excessive degradation of the substrate KRT14 to impair skin integrity.
OK, now we know that the patient's skin lesions were caused by abnormal large-scale degradation of keratin KRT14, and the real magic hand behind this is due to the KLHL24 mutation.
Since KLHL24 is so important and traces its origins, will it also participate in the differentiation process of keratinocytes?
To investigate the regulatory role of KLHL24 in skin differentiation, the authors used a calcium switch to induce a mouse keratinocyte differentiation model. The study found:
Klhl24-encoded mRNA is greatly increased during differentiation, to some extent similar to keratinocyte differentiation markers Lor, Krt10.
The Krt14 protein content decreased significantly with differentiation, while its mRNA level increased slightly, indicating that protein degradation occurred during this process.
According to previous studies, the expression of Klhl24 may be involved in the transcription factor p73, and p73 may be up-regulated in keratinocyte differentiation.
By FISH, it was found that basal keratinocytes expressed lower levels of KLHL24 than basal keratinocytes in normal and patient-derived skin samples, whereas KRT14 was reversed (Fig. 6).
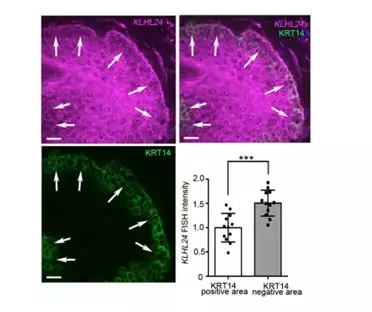
Figure 6. KLHL24 mRNA FISH experiment in skin samples of Patient 5: KLHL24 mRNA FISH (Magenta), KRT14 IF (Green). Arrows: upper periphery of the basal layer. Dot plot: quantification of the FISH intensity of the keratinocytes' cytoplasmic area With positive or negative KRT14 staining respectively.
In conclusion, it was shown that during the differentiation of keratinocytes, KLHL24 expression was up-regulated and played a role in clearing KRT14.
The in vitro study came to an end and it was time to go to the living mouse model.
Construction of heterozygous (Klhl24 c.3G/T ) and homozygous (Klhl24 -/- ) gene-editing mice using CRISPR/Cas9 technology (Fig. 7d) (This model mouse is supplied by Bio-Study and can be used in the Nature Journal It’s also a privilege, honoured. )
Morphological aspects:
Compared with wild-type littermates, Klhl24 c.3G/T mice showed more obvious age-related alopecia, weight loss and small body symptoms (Fig. 7e), but Klhl24 -/- was normal. . None of the heterozygous (Klhl24 c.3G/T ) and homozygous (Klhl24 -/- ) mice exhibited skin fragility.
Protein expression:
Klhl24 c.3G/T , Klhl24 -/- mice and wild-type mouse skin and brain tissues were extracted for immunoprecipitation and immunoblotting experiments. No bands were found in the skin tissue, and Klhl24-∆N28 bands were detected in Klhl24 c.3G/T in brain tissue (Fig. 7f). These results are consistent with the findings of Human Protein Atlas, which means that Klhl24 is expressed more in brain tissue than in skin tissue.
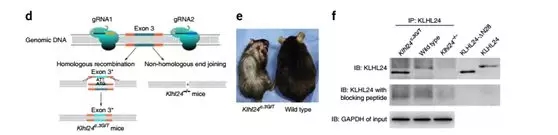
Fig. (d) Illustration of the CRISPR/Cas9-mediated knock-in and knockout strategies. (e) Photograph of a 20-weekold Klhl24c.3G/T male mouse (left) and its wild-type male littermate (right). (f) Cerebral tissue extracts from a Klhl24c.3G/T mouse, its wild-type littermate and a Klhl24-/- mouse were immunoprecipitated with an antibody to KLHL24 followed by immunoblotting.
At the same time, Krt14 content in Klhl24 c.3G/T mouse skin samples was lower than that in wild type (Fig. 7g–i), but the degree of reduction was significant but still higher than Krt14 content in patients' skin. This phenomenon explained Klhl24 c to some extent. .3G/T mice have no skin fragile phenotype.
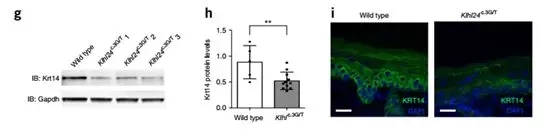
Fig. (g) Immunoblotting using skin extracts shows a reduction in Krt14 protein levels in Klhl24c.3G/T mice in comparison to a wild-type littermate. (h) Quantification of the relative intensity of the Krt14 bands for Klhl24c.3G/T Mic (n = 5) and their wild-type littermates (n = 11) from five independent experiments. (i) Immunofluorescence of Krt14 in skin sections from a Klhl24c.3G/T mouse and its wild-type littermate.
Focus
KLHL24 can be used as an E3 ubiquitin ligase to ubiquitinate the substrate KRT14 and found that its acquired functional mutants can cause a significant decrease in KRT14 and trigger a skin fragility disorder in humans (Fig. 8).
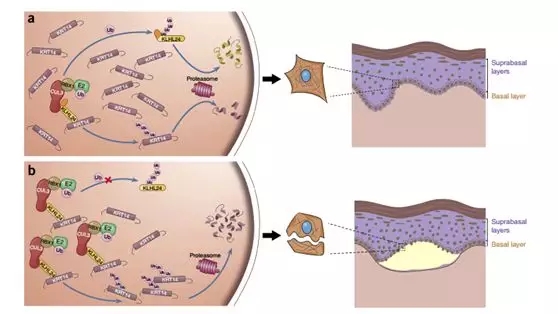
Figure 8. Schematic of the disease-causing mechanism of KLHL24 mutations. (a) Under normal condition, KLHL24 protein is tightly controlled by autoubiquitination and turnover of KRT14 by CUL3–RBX1–KLHL24 is at a low level. (b) In the EBS Case we found, KLHL24-∆N28 is stabilized through abolished autoubiquitination. Overstabilized mutant KLHL24 leads to excessive ubiquitination and degradation of KRT14.
In the differentiation of keratinocytes, the induced expression of KLHL24 and its ubiquitination of KRT14 may be involved, and the self-ubiquitination of KLHL24 can prevent the excessive decrease of KRT14 , thus providing a layer of safety.
In terms of disease phenotype of EB, there will be some differences between human and corresponding mouse models; in our study, mutations in the KLHL24 initiation codon result in different degrees of KRT14 content, as well as differences in humans and mice. The skin disease phenotype, as to why this difference occurs, remains to be further studied.
Conclusion
Losing the N-terminal 28 amino acids can cause KLHL24 to madly degrade the skin structural protein KRT14, which in turn causes skin lesions in humans. So, next, how to solve the problem, how to study other members of the E3 ubiquitin ligase family, is there a little bit of thinking? There is not much to say here, Xiaobian, everyone is interested in reading the literature by themselves~
references:
Feng, C., Lin, Z., Li, S., Wang, H., Yang, Y., & Tan, X. (2017). 474 Stabilizing mutations of KLHL24 ubiquitin ligase cause loss of keratin 14 and human skin fragility Journal of Investigative Dermatology, 137(5), S82.

Scan code to pay attention to the 100 Olympics map to learn more about consulting
Ultrasonic Spot Welding Machine
Product categories of Plastic Ultrasonic Spot Welding Machine, we are specialized manufacturers from China,Portable Ultrasonic Plastic Welding Machine, Digital Ultrasonic Spot Welder suppliers/factory, wholesale high-quality machine of ultrasonic Spot welding R & D and manufacturing, we have the perfect after-sales service and technical support. Look forward to your cooperation!
Ultrasonic Spot Welding Machine,Portable Ultrasonic Plastic Welding Machine,Ultrasonic Spot Welding Machine For Mask,Digital Ultrasonic Spot Welder
Wuxi DIZO Ultrasonic Technology Company , https://www.dizosonic.com