Introduction
Ca v 1.3 is an L-type voltage-gated calcium channel and an important therapeutic target for drug discovery. Studies have shown that some of the drugs on the Ca v 1.3 channel produced a state-dependent effect of 1, which means that the effects of drugs with the changes in membrane potential (Vm) and the consequent changes in the channel status (open, closed Changed and inactivated. Since this may provide highly desirable selectivity for pathologically over-activated Ca v 1.3 channels, there is an increasing need to develop high-throughput analytical methods to assess the effects of channel blockers in different states.
The current screening method uses electrophysiology or fluorescence, using potassium ions to regulate membrane potential, but both methods have significant limitations. Electrophysiology directly controls Vm, but it is invasive and not well suited for high-throughput assays, while fluorometry has poor physiological relevance and is irreversible.
To address these limitations, we report here a new solution that uses optogenetics to control Vm via channel-sensitive channels-2 (channelrhodopsin-2, ChR2) 2 . ChR2 is a non-selective cation channel for light-gating mechanisms and is primarily used to penetrate sodium ions (Na+). When activated by blue light (470 nm), ChR2 mediates the influx of Na + , thereby prolonging the depolarization of the membrane, causing the Ca v 1.3 channel to first enter the open state and then gradually enter the inactive state. Upon termination of photoactivation, the membrane potential is repolarized, which will return the Ca v 1.3 channel to its resting state. In this study, we will use the FLIPR system to demonstrate a new application of optogenetics to control Vm and to reversibly and accurately screen state-dependent calcium channel blockers.
material
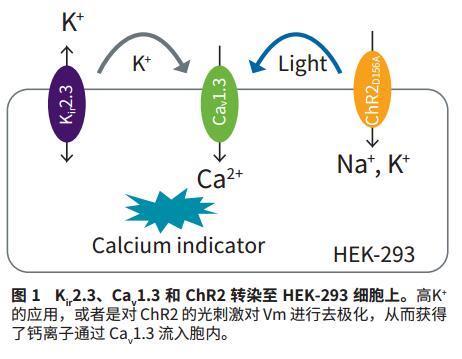
- HEK-293 cell line combined with expression of ChR2D156A (derived from Chlamydomonas reinhardtii), human Ca v 1.3α1 + α2δ, Β subunit, and human K ir 2.3 were supplied by AxxamS.pA, Milan, Italy. The expression of K ir 2.3 first sets Vm to a hyperpolarized value, which can be performed on extracellular high potassium ions (acting on K ir 2.3) or blue light-stimulated ChR2 in the presence of all-trans retinal cofactors. Depolarization. In both cases, depolarized Vm will cause Ca v 1.3 to open, allowing calcium to enter the cell.
- FLIPR TETRA System (0310-5147), FLIPR ® Calcium 6 Assay Kit (R8190) and FLIPR ® Membrane Potential Detection Kit (R8042), all from Molecular Devices.
Advantage
- Develop targeted and non-invasive detection methods through the application of light.
- Flexible, reversible inspection solution
- Data quality meets standards for electrophysiological and potassium ion stimulation methods
method
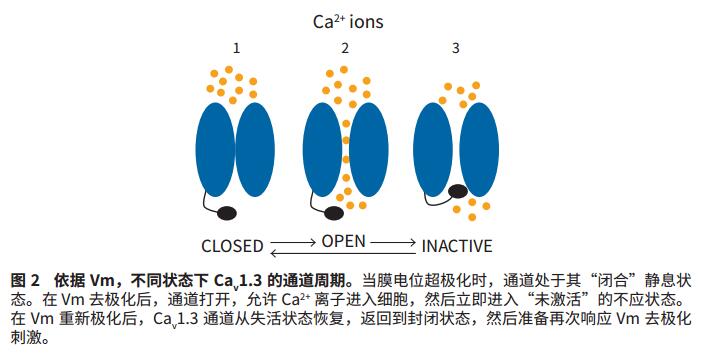
A powerful application of optogenetics is the use of photosensitive drivers to control Vm. In the present study, C. reinhardtii's light-sensitive ChR2 protein was used to control Vm as the basis for high-throughput assays for state-dependent Ca v 1.3 blockers. Specifically, the ChR2 D156A mutant was activated on the FLIPR Tetra system by blue light (470 nm) emitted by its LED, which resulted in an extended time constant of approximately 6.9 minutes due to its permeability to Na + and open state 3 . This results in a super strong and extended membrane depolarization. Due to the depolarization of Vm, the Ca v 1.3 channel first opens allowing Ca 2+ ions to enter the cell (Fig. 1) and enters the inactive state over time (Fig. 2). Dynamic changes in intracellular calcium ions were observed using the FLIPR Calcium 6 Assay Kit.
Control Vm with ChR2
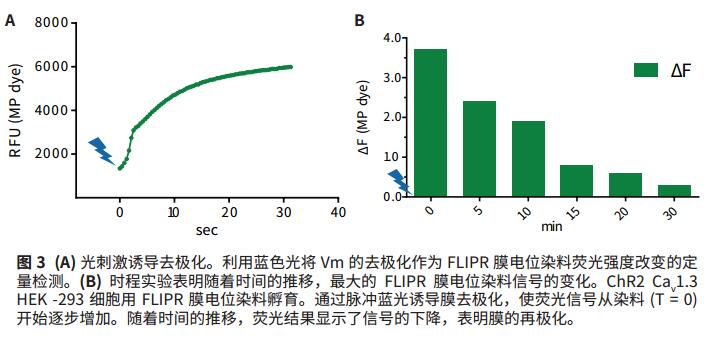
As a first step in verifying Vm control, the change in fluorescence signal was measured with a FLIPR membrane potential dye (Ex530nm/Em565nm), which is a result of the change in Vm. HEK-293 cells were transfected with ChR2, K ir 2.3, and Ca v 1.3 and incubated in the presence of all-trans retinal cofactors according to standard protocol FLIPR membrane potential dyes. Using the FLIPR TETRA system, the light pulse of the blue LED is used to stimulate the ChR2 channel to induce membrane depolarization. The second option is simultaneous, which stimulates the membrane potential dye and measures changes in fluorescence emission over time. The increase in fluorescence signal reflects the depolarization of the membrane. After the first blue light stimulus is over, the ChR2 channel enters a closed state, allowing Vm to repolarize, resulting in a decrease in fluorescence signal over time.
ChR2 control of Ca v 1.3
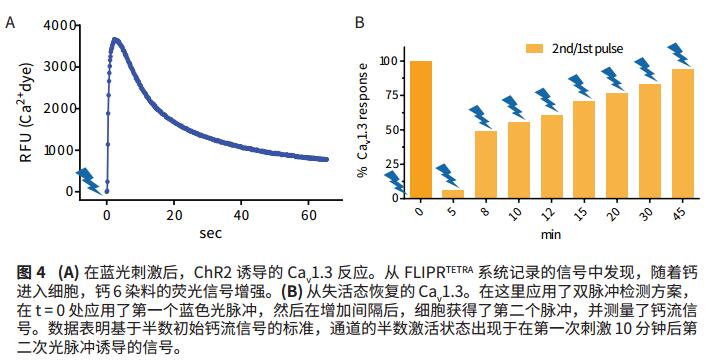
HEK-293 cells were transfected with ChR2, K ir 2.3, and Ca v 1.3, firstly in the presence of all-trans retinal cofactors, and FLIPR calcium 6 dye was added for incubation. Using the FLIPR Tetra system, cells stimulated ChR2 with a blue light pulse, while changes in fluorescence reflect calcium entry into the cell through Ca v 1.3. In the subsequent experiments, a dual pulse stimulation protocol was used, after the first pulse, the second light pulse was applied to the inactivated Ca v 1.3 recovery after the first pulse.
Regulation of Ca v 1.3 under high potassium conditions
For reference, a standard method for analyzing Ca v 1.3 channel activity under high potassium conditions was performed on the same cell line. When high potassium ions are added to the extracellular fluid, membrane potential depolarization causes the Ca v 1.3 channel to open and calcium ions to enter the cell (Figure 1). Over time, Ca v 1.3 enters an inactive, non-conductive state. The increase in intracellular calcium was measured using FLIPR Calcium 6 dye. This method requires that high concentrations of K+ have many "non-targeting" effects on non-physiological cells. Moreover, this application is irreversible because potassium ions cannot be removed from the orifice.
Pharmacology
To validate the utility of optogenetics in pharmacological experiments, a set of experiments used the state-dependent inhibitor erradipine to block the Ca v 1.3 channel and analyze the IC 50 results in the off and inactivated state. Three methods activated Ca v 1.3 channels, including optogenetics, K ir 2.3 based high potassium addition, and patch clamp detection methods, comparing the IC 50 values ​​of eraradipine in both states.
result
Regulating Vm by ChR2
The regulation of membrane potential by photoactivated ChR2 was verified by incubating ChR2-transfected HEK-293 cells with FLIPR membrane potential dye. The blue light pulse emitted by the FLIPR TETRA system LED causes the membrane to be depolarized, while the FLIPR membrane potential dye signal is increased by a factor of three (Fig. 3a). Over time, recording of the membrane potential signal showed that when the cell membrane was repolarized, the fluorescent signal returned to the background level from the initial blue light stimulation to about 30 minutes. This data indicates that when blue light stimulates ChR2 to open, Vm is first depolarized, and as time passes, as the ChR2 channel turns off Vm repolarization (Fig. 3b), its time constant is 6.9 minutes.
ChR2-induced Ca v 1.3 response
Transfected HEK-293 cells were incubated with FLIPR Calcium 6 dye. As calcium ions enter the cell (Fig. 4A), the light pulse emitted by the blue LED triggers an increase in the fluorescent signal. In a subsequent experiment (Fig. 4B), cells of the dual pulse detection protocol were used to identify the time dependence of the Ca v 1.3 channel recovery from inactivation. After a 10 minute interval, a signal of half of the original calcium flow was recorded. After the 45 minute interval, almost 100% of the signal was recorded. The data shows that after 10 minutes of the first light pulse, half of the channels are still inactive, resulting in a 50% maximum calcium signal.
Regulation of potassium ions
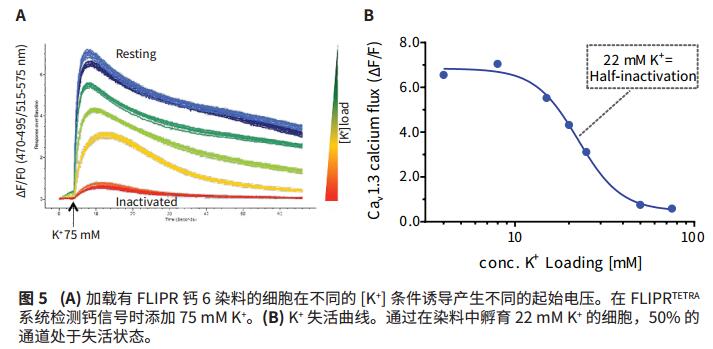
The state of Ca v 1.3 channel was regulated by K + , and the transfected HEK-293 cell line was incubated with FLIPR calcium 6 for one hour at 37 ° C. The buffer was added to increase the potassium ion concentration from 4 mM to 75 mM to form different membrane potentials. (Vm). It was then added to the high concentration K + 75mM calcium detection signal FLIPR TETRA system. A representative calcium signal response is shown in Figure 5A. The K+-dependent inactivation curve shows the FLIPR Calcium 6 dye signal corresponding to the K + concentration, and the data shows that the potassium ion concentration of the induced 50% max Ca2 + signal is 22 mM (Fig. 5B), indicating that this concentration was added. Half of the Ca v 1.3 channels were inactivated under potassium ion conditions.
Pharmacology
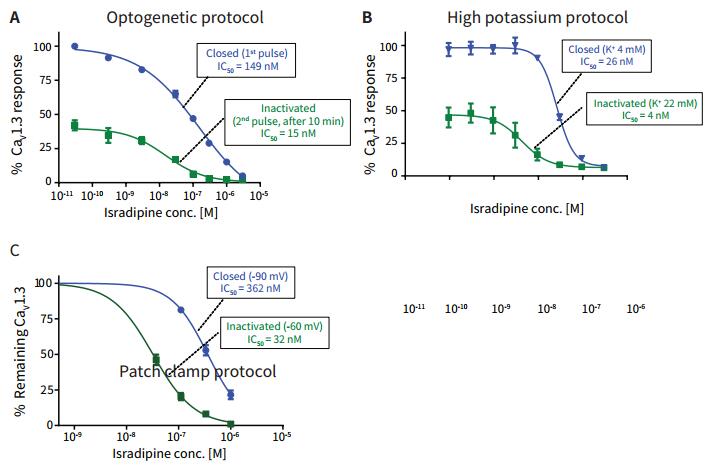
Iradipine is an antihypertensive drug that may also help slow the progression of Parkinson's disease symptoms and was selected as the subject of this study because of its state-dependent effects on Ca v 1.3. Using the optogenetic assay of ChR2, HEK-293 cells expressing Ca v 1.3 were incubated with FLIPR Calcium 6 dye for one hour at 37 ° C with the addition of different concentrations of erradipine. Stimulation was performed using a blue light pulse on the FLIPR TETRA system to detect an increase in the intensity of the calcium flow fluorescent signal in the closed state of the channel. After 10 minutes, a second blue light pulse stimulation was given, and the change in the calcium flow fluorescence signal in the channel inactivation state was recorded. Only half of the intensity of the original calcium flow signal was observed. In Figure 6A, the results are shown as % of the Ca v 1.3 channel effect. High K + regulatory programs recorded isradipine the IC 50 curve shown in Figure 6B. Based on an earlier potassium ion regulation experiment, 4 mM K + or 22 mM K + was added to the loading buffer of FLIPR Calcium 6 dye to induce closure and inactivation, respectively. 75 mM K + was added to detect calcium signals in the FLIPR TETRA system. The IC 50 curve of eraradipine in the Ca v 1.3 channel closed and inactivated state detected under the patch clamp method is shown in Fig. 6C.

to sum up
The IC 50 values ​​for all of the Ca v 1.3 channels in the closed and inactivated state, including the patch clamp results, are shown in Table 1. The left shift of the IC 50 value in the deactivated state obtained under all detection methods is about 10 times as compared with the value in the off state. This is important because the combined effects on ion channels may have different clinical implications in channel closure and inactivation. Light by genetic testing program, IC 50 values closer to the IC 50 values obtained by electrophysiological methods. The "blocking ratio" value obtained by optogenetics is highly similar to that obtained by electrophysiological methods, suggesting a high biological correlation. Unlike electrophysiology, cells in optogenetics are intact at the end of the experiment. In addition, light-driven optogenetic detection is reversible, which is a highly desirable feature of flexible analytical methods.

Acknowledgement
Experimental work and data were provided by AxxamS.pA, Milan, Italy.
references
1. Koschak, A., et al, J. Biol. Chem. 2001, 276.
2. Prigge, M., Rossler, A., Heggeman, P., Channels. 2010, May/June, 4:3, 241-247.
3. Berndt, A., Yizhar, O., Gunaydin, L., Hegemann, P., Deisseroth, K., Nat Neurosci. 2009, Feb; 12(2): 229-234.
4. Catterall WA, Perez-Reyes E, Snutch TP, Striessnig J. Pharmacol Rev. 2005 Dec; 57(4): 411-25.
Walking Frame,Walking Aid Frame,Aluminum Walking Frame,Walking Support Frame
XIAMEN DORRELLA HEALTH TECHNOLOGY CO.,LTD , https://www.dinglimed.com